Metabolická specifičnost cvičení a tréninku je založena na porozumění přenosu energie v biologických soustavách. Účinný a produktivní tréninkový program je možno vytvořit pouze díky porozumění procesu, jakým se vytváří energie pro konkrétní typy cvičení a jak lze přenos energie upravovat pro konkrétní tréninkové režimy.
Základní terminologie
Bioenergetika neboli tok energie v biologické soustavě zahrnuje především přeměnu makroživin, jako sacharidů, bílkovin a tuků, které obsahují chemickou energii.
Energie je schopnost nebo kapacita provádět práci. Rozklad vysoce energetických vazeb v těchto makroživinách uvolňuje energii potřebnou k vykonání biologické práce.
Katabolismus je rozklad velkých molekul na menší molekuly, což je spojeno s uvolněním energie (např. rozklad glykogenu na glukózu).
Anabolismus je opak katabolismu. Je to syntéza větších molekul z menších molekul (např. syntéza bílkovin z aminokyselin).
Exogenní reakce jsou reakce, které uvolňují energii a jsou ve většině katabolické (např. rozklad adenosintrifosfátu na adenosindifosfát).
Endogenní reakce vyžadují energii a zahrnují anabolické procesy.
Metabolismus je souhrnem všech katabolických nebo exogenních a anabolických nebo endogenních reakcí v biologickém systému. Energie získaná katabolicky nebo exogenně se využívá k řízení anabolických nebo endogenních reakcí prostřednictvím molekuly adenosintrifosfátu (ATP).
Adenosintrifosfát umožňuje přenos energie z exogenních na endogenní reakce. Bez dostatečné zásoby ATP by nebyla možná svalová činnost ani růst svalů.
Obrázek 4 Molekula Adenosintrifosfátu (ATP)
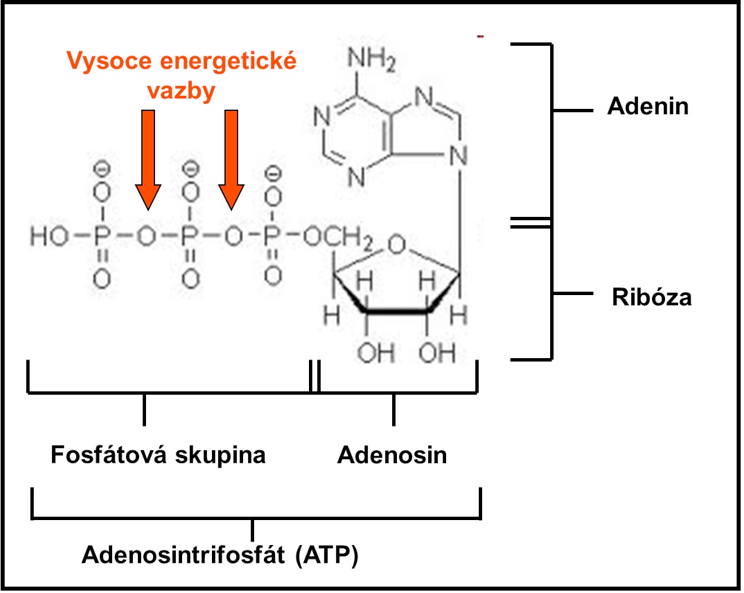
Adenosintrifosfát je tvořen adenosinem a třemi fosfátovými skupinami. Adenosin je sloučenina adeninu (na dusíkovém základu) a ribózy (pěti uhlíkového cukru). Rozklad jedné molekuly ATP k získání energie se nazývá hydrolýza, protože vyžaduje jednu molekulu vody. Hydrolýza ATP je katalyzována přítomností enzymu zvaného adenosintrifosfatáza (ATPáza). Adenosintrifosfát je klasifikován jako vysokoenergetická molekula, protože obsahuje velké množství energie v chemických vazbách dvou koncových fosfátových skupin.
Hydrolýza ATP
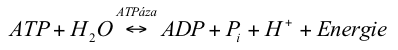
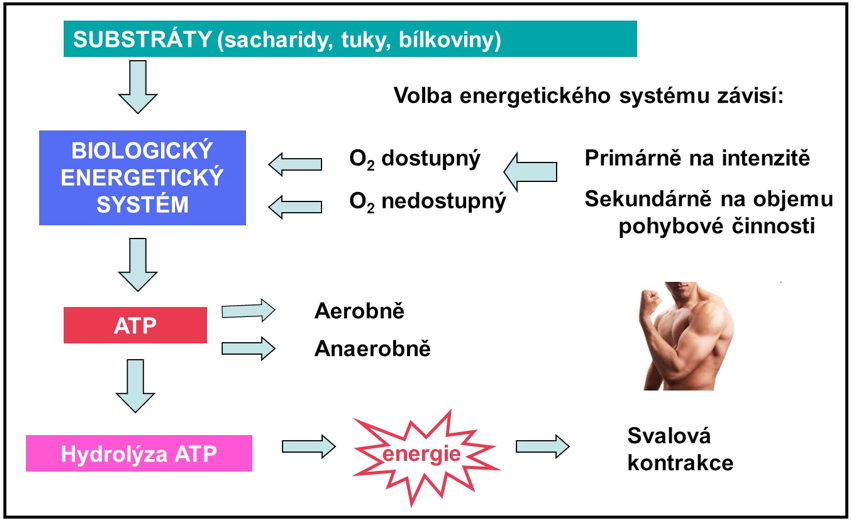
Biologické energetické systémy
V lidských svalových buňkách existují tři energetické systémy k doplnění ATP:
- fosfagenový systém
- glykolýza
- oxidativní systém
Jsou známy dva typy metabolismu (anaerobní a aerobní). Anaerobní procesy nevyžadují přítomnost kyslíku. Fosfagenový systém a první fáze glykolýzy (rychlá glykolýza) jsou anaerobní mechanismy, které probíhají v sarkoplazmě svalových buněk. Krebsův cyklus, přenos elektronů a zbytek oxidačního sytému (pomalá glykolýza, oxidativní systém) jsou aerobní mechanismy, které probíhají v mitochondriích svalových buněk a vyžadují přítomnost kyslíku jako koncového elektronového receptoru.
Fosfagenový systém poskytuje energii velmi krátkou dobu na začátku pohybové činnosti prostřednictvím hydrolýzy zásob ATP a rozkladu CP (kreatin fosfátu).
Rychlá glykolýza využívá sacharidy jako substrát pro tvorbu ATP při činnostech vysoké intenzity bez nutnosti přítomnosti kyslíku. Konečným produktem rychlé glykolýzy je pyruvát, který je dále konvertován na laktát.
Pomalá glykolýza využívá sacharidy jako substrát pro tvorbu ATP při činnostech střední a mírné intenzity, kde konečný produkt glykolýzy pyruvát není konvertován na laktát, ale je transportován do mitochondrií, kde podstoupí Krebsův cyklus. Nutnou podmínkou pomalé glykolýzy představuje dostatečné množství přítomného kyslíku.
Oxidativní systém využívá tuky jako substrát pro tvorbu ATP při činnostech mírné intenzity, kde tuky vstupují přímo do Krebsova cyklu za předpokladu dostatečného množství přítomného kyslíku.
Ze tří hlavních makroživin (sacharidů, bílkovin a tuků) jsou to pouze sacharidy, které mohou být metabolizovány pro energii bez přímého zapojení kyslíku. Proto jsou sacharidy kritické během anaerobního metabolismu. Všechny tři energetické systémy jsou aktivní v jakoukoli danou dobu. Míra, do jaké systémy přispívají k celkovému pracovnímu výkonu, závisí primárně na intenzitě činnosti a sekundárně na délce trvání.
Obrázek 5 Princip úhrady energie v lidském těle
Vysvětlivky: ATP - Adenosintrifosfát
Fosfagenový systém
Fosfagenový systém zajišťuje ATP primárně pro krátkodobé činnosti s velkou intenzitou (např. silový trénink nebo sprint) a je aktivní na začátku všech cvičení bez ohledu na intenzitu. Tento energetický systém spoléhá na hydrolýzu ATP a na rozklad další vysokoenergetické molekuly fosfátu zvané kreatinfosfát (CP). Kreatinkináza je enzym, který katalyzuje syntézu AT z CP a ADP následující reakcí:
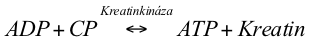
Kreatinfosfát dodává fosfátovou skupinu, která se slučuje s ADP a doplňuje ATP. Kreatinkinázová reakce zajišťuje energii na vysoké úrovni. CP se ukládá v relativně malém množství. Fosfagenový systém nemůže být primárním dodavatelem energie pro nepřetržité, déletrvající činnosti.
ATP je v těle uložen v množství cca 80 až 100 g v jakoukoli danou dobu, což nepředstavuje pro cvičení žádnou významnější zásobu. Koncetrace CP v kosterním svalstvu je čtyři až šestkrát vyšší než koncentrace ATP. Proto slouží fosfagenový systém, za pomoci CP a reakce kreatinkinázy, jako zásoba energie pro rychlé doplnění ATP. Mimo to svalová vlákna typu II (rychlá vlákna) obsahují vyšší koncentraci CP než vlákna typu I (pomalá).
Další důležitá enzymatická reakce, která může rychle doplnit ATP, je adenylátkinázová (také nazývaná myokinázová) reakce:

Tato reakce je obzvláště důležitá, protože AMP, produkt adenylátkinázové (myokinázové) reakce, je účinným stimulátorem glykolýzy.
Řízení fosfagenového systému
Reakce fosfagenového systému jsou ve velké míře řízeny zákonem hromadné akce. Zákon hromadné akce říká, že koncentrace reaktantů nebo produktů (nebo obou) v roztoku bude řídit směr reakce. Například s tím, jak je ATP hydrolyzovaná k získání energie nezbytné pro cvičení, dochází k přechodnému zvýšení koncentrace ADP v sarkolemě. To zvýší poměr kreatinkinázy a adenylátkinázové reakce k doplnění zásoby ATP.
Glykolýza
Glykolýza znamená rozklad sacharidů – buď glykogenu uloženého ve svalu, nebo glukózy dodané do krve – k resyntéze ATP. Proces glykolýzy zahrnuje vícečetné enzymaticky katalyzované reakce. Výsledkem je, že rychlost resyntézy ATP během glykolýzy není rychlá, jak je tomu u fosfagenového systému, avšak její množství je mnohem větší kvůli většímu zásobení glykogenem a glukózou ve srovnání s CP. Jako u fosfagenového systému se objevuje v sarkoplazmě.
Konečným výsledkem glykolýzy je pyruvát, který může pokračovat jedním ze dvou následujících směrů:
- pyruvát může být přeměněn na laktát
- pyruvát může být přesunut do mitochondrií
Když je pyruvát přeměněn na laktát, resytéza ATP probíhá rychleji a závisí na intenzitě a době trvání pohybové činnosti. Tento proces se nazývá anaerobní glykolýza (rychlá glykolýza). Když je pyruvát přesunut do mitochondrií, aby podstoupil Krebsův cyklus, rychlost resyntézy ATP je pomalejší, ale může trvat delší dobu, pokud je intenzita cvičení střední. Tento proces se často nazývá aerobní glykolýza (pomalá glykolýza). Jelikož glykolýza sama o sobě nezávisí na kyslíku, užití pojmů anaerobní a aerobní glykolýza není pravděpodobně příliš praktické pro popis těchto procesů. Potřeba energie závisí primárně na intenzitě pohybové činnosti. Jestliže energie musí být rychle použita, jako například během silového tréninku, pyruvát se primárně přemění na laktát. Jestliže potřeba energie není tak velká a v buňce je v dostatečném množství přítomen kyslík, pyruvát může být dále oxidován v mitochondrii, jako například při vytrvalostním tréninku.
Tvorba laktátu
Tvorba laktátu z pyruvátu se katalyzuje enzymem laktát dehydrogenáza. Běžné množství koncentrace laktátu v krvi je v rozsahu od 0,5 do 2,2 mmol/L v klidu a 0,5 až 2,2 mmol na každý kilogram aktivního svalu. Tvorba laktátu se zvyšuje s intenzitou cvičení a zdá se, že závisí na typu svalového vlákna. Vyšší tvorba laktátu svalovými vlákny typu II může odrážet vyšší koncentraci nebo činnost glykolických enzymů než u svalového vlákna typu I. Celková únava se může vyskytnout při krevní koncentraci mezi 20 a 25 mmol/L. Nejvyšší krevní koncentrace se objevuje zhruba 5 minut po ukončení cvičení.
Akumulace laktátu v krvi je větší po intervalovém zatížení větší intenzity (např. silový trénink a sprint) než po nepřerušovaném zatížení menší intenzity. Trénovaní sportovci prokazují v krvi nižší koncentraci laktátu než netrénovaní jedinci při stejné velikosti zatížení při silovém tréninku.
Koncentrace laktátu v krvi odráží produkci laktátu a jeho odbourání. Laktát lze odbourat oxidací v rámci svalového vlákna, v němž byl vytvořen, nebo může být přenesen krví do jiného svalového vlákna, kde je následně oxidován. Laktát může být také přenesen krví do jater, kde se přemění na glukózu. Tento proces se nazývá Coriho cyklus.
Mezní hodnota laktátu a akumulace krevního laktátu
V literatuře je potvrzeno, že existují konkrétní body zlomu na křivce akumulace laktátu v závislosti na tom, jak se zvyšuje intenzita cvičení. Intenzita cvičení nebo relativní intenzita, při které krevní laktát začíná rapidně narůstat nad hranici základní koncentrace, se nazývá mezní hodnota laktátu (MHL). MHL představuje zvýšený nárok pohybové aktivity na anaerobní mechanismy. MHL velmi dobře koresponduje s ventilačním prahem (VP). Jedná se o bod zlomu ve vztahu mezi ventilací a VO2. V praxi často používán pro odhad anaerobního prahu (viz. kapitola 5).
MHL zpravidla začíná mezi 50% až 60% maximální spotřeby kyslíku u netrénovaných jedinců a mezi 70% až 80% u trénovaných sportovců. Uvedené hodnoty jsou pouze orientační, důvodem jsou velké interindividuální rozdíly u jednotlivých sportovců. V praxi zpravidla nastává výraznější nárůst laktátu při koncentraci mezi 2 a 8 mmol/L krve. Tento rozsah koncentrace laktátu se nazývá Maximální laktátový setrvalý stav (MLSS). V rozsahu intervalu MLSS je při pohybové činnosti tvorba a utilizace laktátu v rovnováze.
Oxidativní (aerobní) systém
Oxidativní systém, primární zdroj ATP v klidovém stavu a během činností nízké intenzity, vyžívá jako substrát především sacharidy a tuky. Po začátku činnosti, kdy intenzita cvičení narůstá, začínají být jako substrát upřednostňovány místo tuků sacharidy. Během aerobních cvičení s vysokou intenzitou je téměř 100% energie vytvořeno ze sacharidů za předpoklad, že je jich dostatek.
Oxidace glukózy a glykogenu
Oxidativní metabolismus krevní glukózy a svalového glykogenu začíná glykolýzou. Pokud je kyslík přítomen v dostatečném množství, výsledný produkt glykolýzy pyruvát není konvertován na laktát, ale je transportován do mitochondrií, kde vstupuje do Krebsova cyklu (pomalá glykolýza) a následně do elektronového transportního řetězce (ETC) za účelem vytvoření ATP z ADP. Produkce ATP touto cestou se nazývá oxidativní fosforylace.
Oxidace tuků
Oxidativní energetický systém může využívat tuky jako zdroj energie. Triglyceridy uložené v tukových buňkách jsou rozštěpeny prostřednictvím enzymu senzitivní lipáza. To uvolňuje volné mastné kyseliny z tukových buněk do krve, kde obíhají a následně se dostávají do svalových vláken a vstupují přímo do Krebsova cyklu a následně do elektronového transportního řetězce (ETC) za účelem vytvoření ATP z ADP. Tvorba ATP tímto procesem se nazývá rovněž oxidativní fosforylace.
Obrázek 6 Přehled energetických systémů v lidském těle
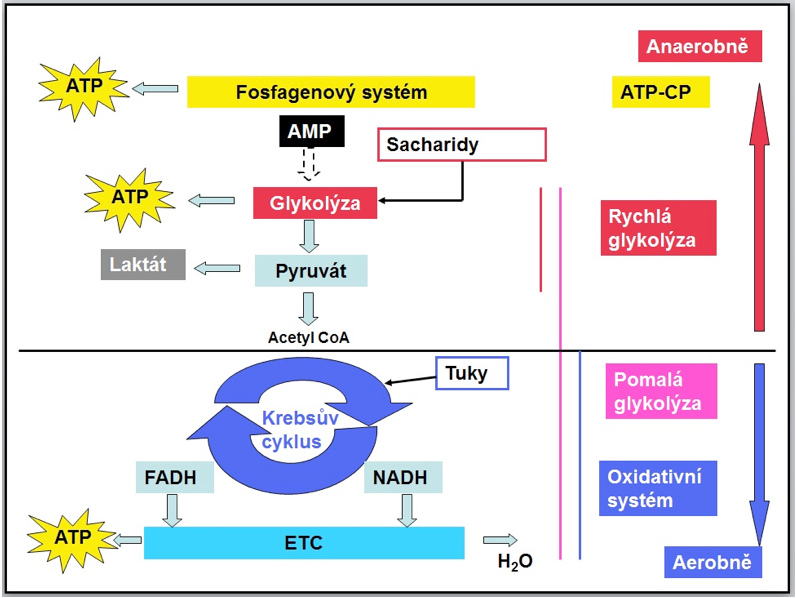
Vysvětlivky: ATP - Adenosintrifosfát, AMP - Adenosinmonofosfát, ATP-CP - Fosfagenový systém, FADH - Flavin adenin dinukleotid, NADH - Nikotinamid adenine dinukleotid, ETC - Elektronový transportní řetězec
Tvorba energie a kapacita
Celková energie získaná oxidací jedné molekuly glukózy je zhruba 40 ATP. Glykolýza spotřebuje buď 2 ATP (jestliže začíná krevní glukózou) nebo 1 ATP (jestliže začíná glykogenem), takže čistá tvorba ATP je 40 – 2 = 38 nebo 40 – 1 = 39. Celková energie získaná z oxidace jedné (18 karbonové) molekuly triglyceridu je 463 ATP. Ostatní triglyceridy, které obsahují různý počet uhlíků, získají více či méně ATP.
Tvorba ATP prostřednictvím výše uvedených energetických systémů se liší svou schopností dodávat energii pro činnosti různé intenzity a trvání. Obecně platí obrácený vztah mezi maximální tvorbou ATP daného energetického systému (tj. ATP vytvořenou za jednotku času) a celkovým množstvím ATP, které je systém schopen vytvořit během delšího časového úseku. Výsledkem je, že fosfagenový energetický systém dodává primárně ATP pro činnosti o vysoké intenzitě a krátkém trvání (např. běh na 100 m), glykolytický systém pro činnosti se střední až velkou intenzitou a krátkým až středně dlouhým trváním (např. běh na 400 m) a oxidativní systém pro činnosti s nízkou intenzitou dlouhého trvání (např. marathon).
Míra, do jaké tyto tři energetické systémy přispívají k tvorbě ATP, záleží primárně na intenzitě svalové aktivity a sekundárně na délce trvání. Nikdy, ať už během cvičení, či odpočinku, nezajišťuje celkový přísun energie pouze jeden samotný systém.
Tabulka 1 Vliv délky trvání činnosti a intenzity na to, který energetický systém bude využit jako primární
| Délka trvání činnosti | Intenzita činnosti | Primární energetický systém (systémy) |
|---|---|---|
| 0-6 sekund | Extrémně velká | Fosfagenový |
| 6-30 sekund | Velmi velká | Fosfagenový a rychlá glykolýza |
| 30 sekund až 2 minuty | Velká | Rychlá glykolýza |
| 2-3 minuty | Střední | Rychlá glykolýza a oxidační systém |
| >3 minuty | Malá | Oxidační systém |
Tabulka 2 Měřítko stupně a kapacity tvorby ATP
| Systém | Stupně tvorby ATP | Kapacita tvorby ATP |
|---|---|---|
| Fosfagenový | 1 | 5 |
| Rychlá glykolýza | 2 | 4 |
| Pomalá glykolýza | 3 | 3 |
| Oxidace sacharidů | 4 | 2 |
| Oxidace tuků a bílkovin | 5 | 1 |
Pozn.: 1 = nejrychlejší/největší; 5 = nejpomalejší/nejmenší
Spotřeba a doplnění substrátu
Energetické substráty jsou molekuly, které zajišťují počáteční materiál pro bioenergetické reakce, včetně fosfagenů (ATP a kreatinfosfát), glukózy, glykogenu, laktátu, volných mastných kyselin a aminokyselin.
Fosfageny
Koncentrace fosfagenů ve svalech se rychle spotřebovává následkem anaerobního cvičení velké intenzity. Kreatin fosfát se může výrazně snížit (50-70%) během krátké doby (5-30s) cvičení o velké intenzitě a může být téměř zcela spotřebován, přičemž výsledkem je vyčerpání po velmi intenzivním cvičení.
Doplnění fosfagenu po cvičení může nastat během relativně krátké doby. Kompletní resyntéza ATP se objevuje do 3 až 5 minut, kompletní resyntéza kreatin fosfátu se objevuje do 8 minut.
Glykogen
Pro cvičení jsou k dispozici jen omezené zásoby glykogenu. Zhruba 300 až 400 g glykogenu je uloženo v celkovém tělesném svalstvu, okolo 70 až 100 g v játrech. Relativně stálá koncentrace krevní glukózy je zachována při cvičeních o velmi malé intenzitě (pod 50% maximální spotřeby kyslíku). Se zvyšující se délkou trvání nad 90 minut se zásoba glukózy zmenšuje, ale pouze zřídka na méně než 2,8 mmol/L. Cvičení probíhající po delší časový úsek (nad 90 minut) při vyšší intenzitě (nad 50% maximální spotřeby kyslíku) může vyústit v soustavně nízkou zásobu glukózy v krvi. Hypoglykemická reakce se může objevit u některých lidí, jimž v důsledku cvičení klesla hladina glukózy pod 2,5 mmol/L. Po ukončení cvičení musí dojít k doplnění sacharidů. Doplnění glykogenu (glykogen je sloučenina molekul glukózy) během odpočinku je spojeno s přijímáním sacharidů po cvičení. Doplnění je optimální, když je každé dvě hodiny po cvičení přijato 0,7 až 3,0 g sacharidů na kilogram tělesné váhy. Svalový glykogen je možno zcela doplnit do 24 hodin za předpokladu, že je přijímáno dostatečné množství sacharidů.
Tuky
Zdroj energie pro déle trvající činnost. V lidském těle zásoba 5-20kg v podkožním tuku, která teoreticky vystačí pro nekonečně dlouhou činnost.
Metabolická specifičnost tréninku
Přiměřená intenzita cvičení a délky odpočinku mohou umožnit „výběr“ konkrétního energetického systému během tréninku pro konkrétní sportovní činnost. Tabulka 3 ukazuje doporučenou délku odpočinku při tréninku konkrétního energetického systému.
Tab. 3 Intenzita, interval zatížení a délka odpočinku pro trénink konkrétního energetického systému